Cell wall
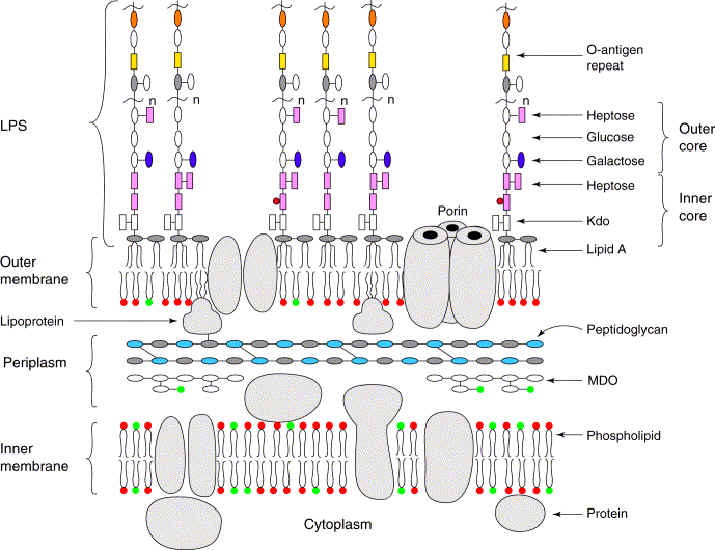
The Gram stain for bacteria allows differentiation according to thickness of the layer of peptidoglycan (murein, im) in the cell wall. Bacteria that stain heavily (Gram +ve) have a thick monolayer of peptidoglycan compared to the thin or absent layer of peptidoglycan in (bilayer) bacteria that do not take up the stain (Gram -ve). The cell walls of Archaeobacteria contain no peptidoglycan (murein), rather they contain pseudomurein, complex carbohydrates, or protein-glycoproteins.
S-layers comprise one of the most common surface structures on archaea and bacteria. These surface layers have now been identified in hundreds of different species belonging to all major phylogenetic groups of bacteria, and they represent a feature common to almost all archaea (recent compilation 133). S-layers are monomolecular crystalline arrays of proteinaceous subunits (125, 131, 132).
Tables Cell walls of Prokaryotes Comparisons of Eubacteria, Archaea, and Eukaryotes Electron acceptors for respiration and methanogenesis in prokaryotes Glycolysis in bacteria Lithotrophic prokaryotes Structure of bacteriochlorophylls Comparison of plant and bacterial photosynthesis :
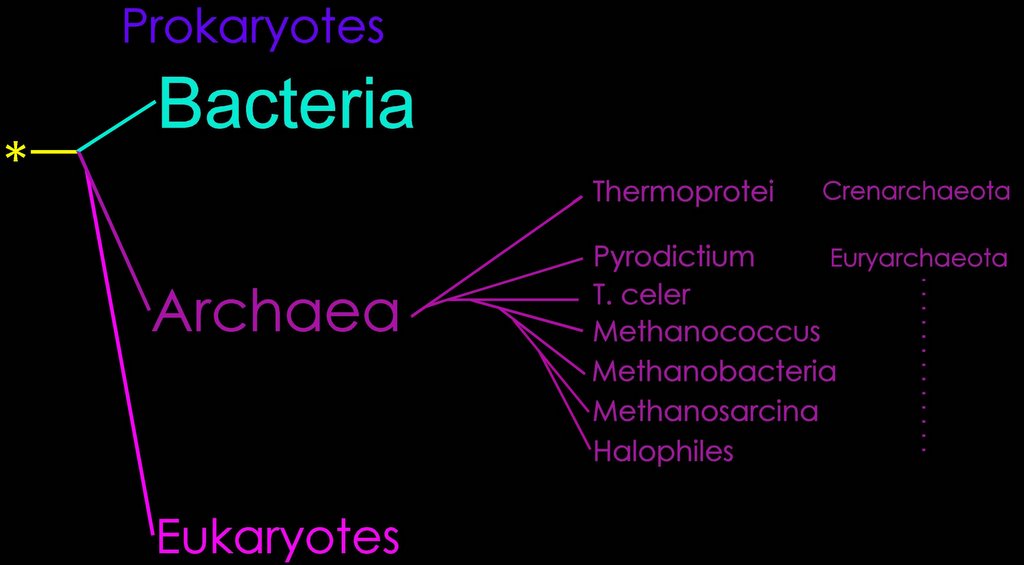
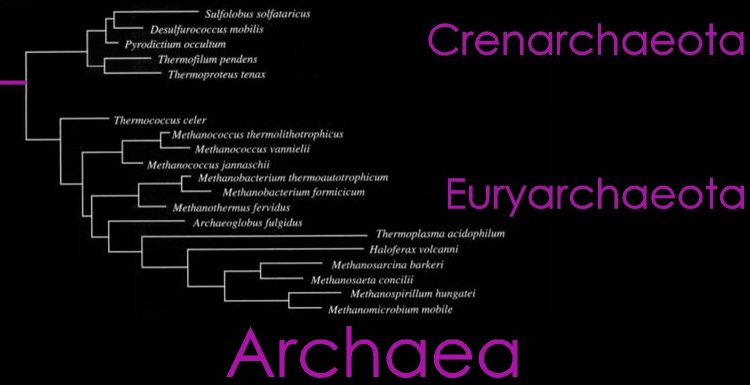
Diagrams: Eubacteria : peptidoglycan : gram + gram - peptidoglycan : Gram + : Gram - : P. aeruginosa comp-mod : gram -ve : antimicrobials gram + gram - : cell walls gram + gram - mycobacteria : comparison gram + / gram - cell walls : b-w g+ g- : Archaea : Gram positive archaeal cell wall : Gram negative archaeal cell wall : Unusual cell wall of Deinococcus radiodurans :
: S-layer Freeze etched sem :
The structure of secondary cell wall polymers: how Gram-positive bacteria stick their cell walls together.
The cell wall of Gram-positive bacteria has been a subject of detailed chemical study over the past five decades. Outside the cytoplasmic membrane of these organisms the fundamental polymer is peptidoglycan (PG), which is responsible for the maintenance of cell shape and osmotic stability. In addition, typical essential cell wall polymers such as teichoic or teichuronic acids are linked to some of the peptidoglycan chains. In this review these compounds are considered as 'classical' cell wall polymers. In the course of recent investigations of bacterial cell surface layers (S-layers) a different class of 'non-classical' secondary cell wall polymers (SCWPs) has been identified, which is involved in anchoring of S-layers to the bacterial cell surface. Comparative analyses have shown considerable differences in chemical composition, overall structure and charge behaviour of these SCWPs. This review discusses the progress that has been made in understanding the structural principles of SCWPs, which may have useful applications in S-layer-based 'supramolecular construction kits' in nanobiotechnology.
Schaffer C, Messner P. The structure of secondary cell wall polymers: how Gram-positive bacteria stick their cell walls together. (Free Full Text Article) Microbiology. 2005 Mar;151(Pt 3):643-51.
Molecular organization of selected prokaryotic S-layer proteins.
Regular crystalline surface layers (S-layers) are widespread among prokaryotes and probably represent the earliest cell wall structures. S-layer genes have been found in approximately 400 different species of the prokaryotic domains bacteria and archaea. S-layers usually consist of a single (glyco-)protein species with molecular masses ranging from about 40 to 200 kDa that form lattices of oblique, tetragonal, or hexagonal architecture. The primary sequences of hyperthermophilic archaeal species exhibit some characteristic signatures. Further adaptations to their specific environments occur by various post-translational modifications, such as linkage of glycans, lipids, phosphate, and sulfate groups to the protein or by proteolytic processing. Specific domains direct the anchoring of the S-layer to the underlying cell wall components and transport across the cytoplasma membrane. In addition to their presumptive original role as protective coats in archaea and bacteria, they have adapted new functions, e.g., as molecular sieves, attachment sites for extracellular enzymes, and virulence factors.
Claus H, Akca E, Debaerdemaeker T, Evrard C, Declercq JP, Harris JR, Schlott B, Konig H.
Molecular organization of selected prokaryotic S-layer proteins. Can J Microbiol. 2005 Sep;51(9):731-43.
Glycoproteins in prokaryotes. [Arch Microbiol. 1997] PMID: 9382700
Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions. [J Struct Biol. 1998] PMID: 10049812
Prokaryotic glycosylation. [Proteomics. 2001] PMID: 11680871
Glycobiology of surface layer proteins. [Biochimie. 2001] PMID: 11522387
Stress genes and proteins in the archaea. [Microbiol Mol Biol Rev. 1999] PMID: 10585970
See all Related Articles...
S-Layer proteins.
Cell walls are an important structural component of prokaryotic organisms and essential for many aspects of their life. Particularly, the diverse structures of the outermost boundary layers strongly reflect adaptations of organisms to specific ecological and environmental conditions (6).
Over the past 3 decades of research, it has become apparent that one of the most common surface structures on archaea and bacteria are monomolecular crystalline arrays of proteinaceous subunits termed surface layers or S-layers (125, 131, 132). Since S-layer-carrying organisms are ubiquitous in the biosphere and because S-layers represent one of the most abundant cellular proteins, it is now obvious that these metabolically expensive products must provide the organisms with an advantage of selection in very different habitats (133). This minireview provides a brief survey of the current state of our knowledge about S-layers with a particular focus on molecular biological and genetic aspects. Other recent reviews (5, 7, 127, 133, 135) are recommended for a more detailed introduction to and treatises on this subject.
Sara M, Sleytr UB. S-Layer proteins. (Free Full Text Article) J Bacteriol. 2000 Feb;182(4):859-68.
Prokaryotic glycosylation. [Proteomics. 2001] PMID: 11680871
Common history at the origin of the position-function correlation in transcriptional regulators in archaea and bacteria. [J Mol Evol. 2001] PMID: 11523004
[Homologous protein domains in superkingdoms Archaea, Bacteria, and Eukaryota and the problem of the origin of eukaryotes] [Izv Akad Nauk Ser Biol. 2005] PMID: 16212260
See all Related Articles...
Cell wall polymers in Archaea (Archaebacteria).
The distribution of the various cell wall and cell envelope (S-layer) polymers among the main lineages of the domain Archaea (Archaebacteria) and the chemical composition and primary structure of polymers forming rigid cell wall sacculi is described. Differences between bacteria and archaea in their sensitivity to antibiotics which inhibit cell wall synthesis in bacteria are discussed.
Kandler O, Konig H. Cell wall polymers in Archaea (Archaebacteria). Cell Mol Life Sci. 1998 Apr;54(4):305-8.
beta-Lactamases are absent from Archaea (archaebacteria). [Microb Drug Resist. 1996] PMID: 9158771
Structure of anionic carbohydrate-containing cell wall polymers in several representatives of the order actinomycetales. [Biochemistry (Mosc). 2000] PMID: 11092967
Anionic polymers in cell walls of gram-positive bacteria. [Biochemistry (Mosc). 1997] PMID: 9360295
The response of selected members of the archaea to the gram stain. [Microbiology. 1996] PMID: 8885405
Life's third domain (Archaea): an established fact or an endangered paradigm? [Theor Popul Biol. 1998] PMID: 9733652
See all Related Articles...
S-layers comprise one of the most common surface structures on archaea and bacteria. These surface layers have now been identified in hundreds of different species belonging to all major phylogenetic groups of bacteria, and they represent a feature common to almost all archaea (recent compilation 133). S-layers are monomolecular crystalline arrays of proteinaceous subunits (125, 131, 132).
Tables Cell walls of Prokaryotes Comparisons of Eubacteria, Archaea, and Eukaryotes Electron acceptors for respiration and methanogenesis in prokaryotes Glycolysis in bacteria Lithotrophic prokaryotes Structure of bacteriochlorophylls Comparison of plant and bacterial photosynthesis :
Diagrams: Eubacteria : peptidoglycan : gram + gram - peptidoglycan : Gram + : Gram - : P. aeruginosa comp-mod : gram -ve : antimicrobials gram + gram - : cell walls gram + gram - mycobacteria : comparison gram + / gram - cell walls : b-w g+ g- : Archaea : Gram positive archaeal cell wall : Gram negative archaeal cell wall : Unusual cell wall of Deinococcus radiodurans :
: S-layer Freeze etched sem :
The structure of secondary cell wall polymers: how Gram-positive bacteria stick their cell walls together.
The cell wall of Gram-positive bacteria has been a subject of detailed chemical study over the past five decades. Outside the cytoplasmic membrane of these organisms the fundamental polymer is peptidoglycan (PG), which is responsible for the maintenance of cell shape and osmotic stability. In addition, typical essential cell wall polymers such as teichoic or teichuronic acids are linked to some of the peptidoglycan chains. In this review these compounds are considered as 'classical' cell wall polymers. In the course of recent investigations of bacterial cell surface layers (S-layers) a different class of 'non-classical' secondary cell wall polymers (SCWPs) has been identified, which is involved in anchoring of S-layers to the bacterial cell surface. Comparative analyses have shown considerable differences in chemical composition, overall structure and charge behaviour of these SCWPs. This review discusses the progress that has been made in understanding the structural principles of SCWPs, which may have useful applications in S-layer-based 'supramolecular construction kits' in nanobiotechnology.
Schaffer C, Messner P. The structure of secondary cell wall polymers: how Gram-positive bacteria stick their cell walls together. (Free Full Text Article) Microbiology. 2005 Mar;151(Pt 3):643-51.
Molecular organization of selected prokaryotic S-layer proteins.
Regular crystalline surface layers (S-layers) are widespread among prokaryotes and probably represent the earliest cell wall structures. S-layer genes have been found in approximately 400 different species of the prokaryotic domains bacteria and archaea. S-layers usually consist of a single (glyco-)protein species with molecular masses ranging from about 40 to 200 kDa that form lattices of oblique, tetragonal, or hexagonal architecture. The primary sequences of hyperthermophilic archaeal species exhibit some characteristic signatures. Further adaptations to their specific environments occur by various post-translational modifications, such as linkage of glycans, lipids, phosphate, and sulfate groups to the protein or by proteolytic processing. Specific domains direct the anchoring of the S-layer to the underlying cell wall components and transport across the cytoplasma membrane. In addition to their presumptive original role as protective coats in archaea and bacteria, they have adapted new functions, e.g., as molecular sieves, attachment sites for extracellular enzymes, and virulence factors.
Claus H, Akca E, Debaerdemaeker T, Evrard C, Declercq JP, Harris JR, Schlott B, Konig H.
Molecular organization of selected prokaryotic S-layer proteins. Can J Microbiol. 2005 Sep;51(9):731-43.
Glycoproteins in prokaryotes. [Arch Microbiol. 1997] PMID: 9382700
Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions. [J Struct Biol. 1998] PMID: 10049812
Prokaryotic glycosylation. [Proteomics. 2001] PMID: 11680871
Glycobiology of surface layer proteins. [Biochimie. 2001] PMID: 11522387
Stress genes and proteins in the archaea. [Microbiol Mol Biol Rev. 1999] PMID: 10585970
See all Related Articles...
S-Layer proteins.
Cell walls are an important structural component of prokaryotic organisms and essential for many aspects of their life. Particularly, the diverse structures of the outermost boundary layers strongly reflect adaptations of organisms to specific ecological and environmental conditions (6).
Over the past 3 decades of research, it has become apparent that one of the most common surface structures on archaea and bacteria are monomolecular crystalline arrays of proteinaceous subunits termed surface layers or S-layers (125, 131, 132). Since S-layer-carrying organisms are ubiquitous in the biosphere and because S-layers represent one of the most abundant cellular proteins, it is now obvious that these metabolically expensive products must provide the organisms with an advantage of selection in very different habitats (133). This minireview provides a brief survey of the current state of our knowledge about S-layers with a particular focus on molecular biological and genetic aspects. Other recent reviews (5, 7, 127, 133, 135) are recommended for a more detailed introduction to and treatises on this subject.
Sara M, Sleytr UB. S-Layer proteins. (Free Full Text Article) J Bacteriol. 2000 Feb;182(4):859-68.
Prokaryotic glycosylation. [Proteomics. 2001] PMID: 11680871
Common history at the origin of the position-function correlation in transcriptional regulators in archaea and bacteria. [J Mol Evol. 2001] PMID: 11523004
[Homologous protein domains in superkingdoms Archaea, Bacteria, and Eukaryota and the problem of the origin of eukaryotes] [Izv Akad Nauk Ser Biol. 2005] PMID: 16212260
See all Related Articles...
Cell wall polymers in Archaea (Archaebacteria).
The distribution of the various cell wall and cell envelope (S-layer) polymers among the main lineages of the domain Archaea (Archaebacteria) and the chemical composition and primary structure of polymers forming rigid cell wall sacculi is described. Differences between bacteria and archaea in their sensitivity to antibiotics which inhibit cell wall synthesis in bacteria are discussed.
Kandler O, Konig H. Cell wall polymers in Archaea (Archaebacteria). Cell Mol Life Sci. 1998 Apr;54(4):305-8.
beta-Lactamases are absent from Archaea (archaebacteria). [Microb Drug Resist. 1996] PMID: 9158771
Structure of anionic carbohydrate-containing cell wall polymers in several representatives of the order actinomycetales. [Biochemistry (Mosc). 2000] PMID: 11092967
Anionic polymers in cell walls of gram-positive bacteria. [Biochemistry (Mosc). 1997] PMID: 9360295
The response of selected members of the archaea to the gram stain. [Microbiology. 1996] PMID: 8885405
Life's third domain (Archaea): an established fact or an endangered paradigm? [Theor Popul Biol. 1998] PMID: 9733652
See all Related Articles...
Labels: Archaea, cell walls, Eubacteria, glycoproteins, Gram stain, peptidoglycan, PG, pseudomurein, S-layers, SCWP, secondary cell wall polymers, teichoic, teichuronic








































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}