Interactions in Bacteria
Quorum sensing employs chemical signals for bacterial communication concerning achievement of critical mass. This process employs the production, release, and subsequent detection of chemical signaling molecules called autoinducers, and enables bacteria to regulate gene expression in response to changes in cell-population density. In response to quorum signals, bacteria alter in unison processes that are effective only when sufficient cells are available for coordinated activity. As bacterial populations grow, extracellular concentration of autoinducer increases until a threshold is reached at which the population responds with a population-wide alteration in gene expression. The processes controlled by quorum sensing are usually unproductive for an individual bacterium yet effective when undertaken by the group –bioluminescence, secretion of virulence factors, biofilm formation, sporulation, and the exchange of DNA.
Bacteria employ multiple chemical signaling molecules called autoinducers (AIs), one of which, AI-2, enables communication between other bacterial species in the vicinity. A gene called luxS is required for production of AI-2, and hundreds of species of bacteria have the luxS gene and employ AI-2 to communicate, suggesting that bacteria have a universal chemical language for communication between species. The LuxI protein synthesizes an acyl-homoserine lactone autoinducer, and the LuxR protein is responsible for autoinducer binding and subsequent activation of transcription of the luciferase operon. Cell-to-Cell Communication in Bacteria . Say What? Bacterial Conversation-Stoppers .
Cell-Cell Interactions in Bacteria : "One current estimate suggests that more than 99% of the bacteria on earth live as cell masses (Costerton et al., 1995), a condition conducive to cell interactions. As the broader significance of multicellular microbial life has been recognized, the cell interactions that facilitate multicellularity have been revealed."
"Now it is generally accepted that bacteria produce, and respond as groups to, chemical signals and that this interaction can lead to the coordination of group bacterial activities. This phenomenon has become known as quorum sensing (2-4). We also understand that groups of bacteria can form physical structures with unique characteristics, so-called biofilms (5, 6). Quorum sensing and biofilm biology have become very active areas in microbiology, and a large group of investigators is working on these fascinating aspects of bacterial biology, hoping to develop new therapeutic agents to treat associated persistent bacterial infections."
E. Peter Greenberg Bacterial communication and group behavior. J. Clin. Invest. 112:1288-1290 (2003). doi:10.1172/JCI200320099
"Quorum sensing uses signaling molecules, known as autoinducers [molecules that regulate mRNA production for specific genes in response to population density]. These are continuously produced by bacteria and can readily diffuse through the cell membrane. When elevated numbers of bacteria are present in an area, the concentration of autoinducers in the region will be higher. Autoinducer molecules (which include certain peptides and compounds known as homoserine lactones) can interact with specific repressor or activator sequences in DNA. The presence or absence of the autoinducer thus controls the production of mRNA, and therefore protein. These proteins are encoded by dozens of genes, including the genes for biofilm production. Laboratory strains of P. aeruginosa lacking the gene for a specific homoserine lactone will not develop into normal biofilms but pile up into a disorganized heap. From the bacteria's perspective, intracellular signaling has many advantages. Microbes often produce antibiotics that inhibit the growth of competitive species. Intracellular signaling not only brings bacteria together in biofilms, it also regulates the coordinated delivery of high doses of these antibiotics from the denser bacterial population. It also helps bacteria coordinate the release of virulence factors (such as disease-causing toxins) to overcome animal or plant defenses. Signals between bacteria in close proximity, as in a biofilm, also seem to enhance bacterial mating and the acquisition of novel DNA by transformation, both of which increase bacterial diversity." Biofilm Formation and Bacterial Communication.
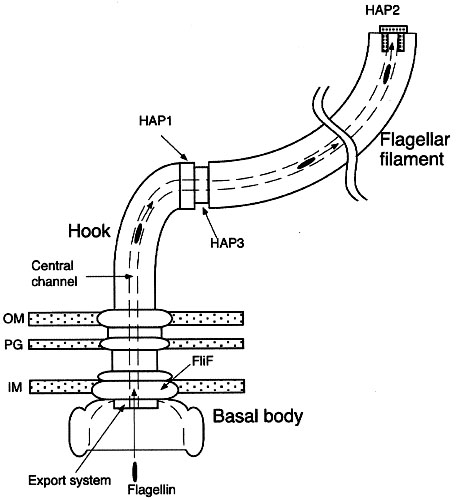
"Now, the Kornberg team has found that P. aeruginosa bacteria without the gene [for PPK, polyphosphate kinase] are also unable to form microbial communities called biofilms, and they are unable to communicate with each other via a process called quorum sensing. Biofilms and quorum sensing are phenomena that occur when the bacteria encounter a new environment and must adapt to survive. Quorum-sensing communication between bacteria involves the release of small molecules that float among the bacteria and deliver a chemical message. When the bacteria determine, via quorum-sensing communication, that a critical mass of organisms has assembled, they use their flagella to slide toward each other and construct a thick, slimy biofilm." Bacterial communication, toxin production tied to intriguing cell protein, and:
"Polyphosphate is a long, chain-like molecule found in every living cell. Scientists believe that in animals, one of its roles may be to serve as a phosphate storage reservoir for the production of ATP (adenosine triphosphate), which provides the energy to power a cell. In bacteria, polyphosphate helps these single-celled organisms adapt to nutritional deficiencies and environmental stresses. It also helps them survive the state of suspended animation known as the stationary phase of growth. "
Bacteria also communicate via conjugation, which enables bacteria to exchange genetic material because of tube-like connections called pili. The donor bacterium contains conjugative or mobilizable genetic elements, usually a conjugative plasmid or episome plasmid that can integrate itself into the bacterial chromosome by genetic recombination. One such conjugative plasmid is called the F-plasmid. This is an episome about 100 thousand base-pairs in length, which carries its own origin of replication, called oriV. Most conjugative plasmids have systems ensuring that the recipient cell does not already contain a similar element, ensuring that there is only one copy of the F-plasmid in the F-positive bacterium. Genetic Construct for F and R cell types.
: New Salmonella Finding—Inter-Bacterial Communication! : Princeton - Bacterial communication :
Mahadevan, S (2002) Bacterial communication. Journal of Biosciences 27(5):pp. 443-444. Full text available as:PDF.
Phosphorylation switches
response regulator proteins in bacteria
Molecular Mechanisms of Signal Transduction : "The majority of bacterial response regulator proteins are transcription factors that serve as repressors or activators to regulate the expression of specific genes. The effector domains of these response regulators are DNA-binding domains that can be categorized into three major families based on sequence and structural similarity.
The OmpR/PhoB family of response regulator transcription factors, distinguished by a winged-helix DNA-binding domain, is the largest family, accounting for ~45 percent of all response regulators. Most characterized members of this family have been shown to bind as tandem dimers to direct repeat DNA recognition sequences. . . Phosphorylation induces dimerization or higher-order oligomerization of the proteins and that dimerization is mediated by the phosphorylated regulatory domains. The activated regulatory domains of Escherichia coli ArcA, KdpE, PhoB, PhoP, and TorR and T. maritima DrrB and DrrD all exist as dimers with identical alpha4-beta5-alpha5 interfaces. In all cases, the dimerization interface is formed by a few hydrophobic residues surrounded by an extensive network of intra- and intermolecular salt bridges. The residues involved in these interactions are highly conserved in all members of the OmpR/PhoB family, but not in other response regulators. It is proposed that this mode of dimerization is common to most members of the OmpR/PhoB family and that it represents a family-specific mechanism for activation of DNA binding. Upon phosphorylation, the interface between regulatory and DNA-binding domains is disrupted, allowing the regulatory domains to dimerize via their alpha4-beta5-aalpha5 faces. Disruption of the interdomain interface frees the DNA-binding domain, allowing it to dimerize in tandem on direct repeat DNA half-sites with symmetry that is different from that of the regulatory domain dimer. Additional nuclear magnetic resonance (NMR) and biochemical studies support this mechanism of activation. " Link to link-to-images.
Bacteria employ multiple chemical signaling molecules called autoinducers (AIs), one of which, AI-2, enables communication between other bacterial species in the vicinity. A gene called luxS is required for production of AI-2, and hundreds of species of bacteria have the luxS gene and employ AI-2 to communicate, suggesting that bacteria have a universal chemical language for communication between species. The LuxI protein synthesizes an acyl-homoserine lactone autoinducer, and the LuxR protein is responsible for autoinducer binding and subsequent activation of transcription of the luciferase operon. Cell-to-Cell Communication in Bacteria . Say What? Bacterial Conversation-Stoppers .
Cell-Cell Interactions in Bacteria : "One current estimate suggests that more than 99% of the bacteria on earth live as cell masses (Costerton et al., 1995), a condition conducive to cell interactions. As the broader significance of multicellular microbial life has been recognized, the cell interactions that facilitate multicellularity have been revealed."
"Now it is generally accepted that bacteria produce, and respond as groups to, chemical signals and that this interaction can lead to the coordination of group bacterial activities. This phenomenon has become known as quorum sensing (2-4). We also understand that groups of bacteria can form physical structures with unique characteristics, so-called biofilms (5, 6). Quorum sensing and biofilm biology have become very active areas in microbiology, and a large group of investigators is working on these fascinating aspects of bacterial biology, hoping to develop new therapeutic agents to treat associated persistent bacterial infections."
E. Peter Greenberg Bacterial communication and group behavior. J. Clin. Invest. 112:1288-1290 (2003). doi:10.1172/JCI200320099
"Quorum sensing uses signaling molecules, known as autoinducers [molecules that regulate mRNA production for specific genes in response to population density]. These are continuously produced by bacteria and can readily diffuse through the cell membrane. When elevated numbers of bacteria are present in an area, the concentration of autoinducers in the region will be higher. Autoinducer molecules (which include certain peptides and compounds known as homoserine lactones) can interact with specific repressor or activator sequences in DNA. The presence or absence of the autoinducer thus controls the production of mRNA, and therefore protein. These proteins are encoded by dozens of genes, including the genes for biofilm production. Laboratory strains of P. aeruginosa lacking the gene for a specific homoserine lactone will not develop into normal biofilms but pile up into a disorganized heap. From the bacteria's perspective, intracellular signaling has many advantages. Microbes often produce antibiotics that inhibit the growth of competitive species. Intracellular signaling not only brings bacteria together in biofilms, it also regulates the coordinated delivery of high doses of these antibiotics from the denser bacterial population. It also helps bacteria coordinate the release of virulence factors (such as disease-causing toxins) to overcome animal or plant defenses. Signals between bacteria in close proximity, as in a biofilm, also seem to enhance bacterial mating and the acquisition of novel DNA by transformation, both of which increase bacterial diversity." Biofilm Formation and Bacterial Communication.
"Now, the Kornberg team has found that P. aeruginosa bacteria without the gene [for PPK, polyphosphate kinase] are also unable to form microbial communities called biofilms, and they are unable to communicate with each other via a process called quorum sensing. Biofilms and quorum sensing are phenomena that occur when the bacteria encounter a new environment and must adapt to survive. Quorum-sensing communication between bacteria involves the release of small molecules that float among the bacteria and deliver a chemical message. When the bacteria determine, via quorum-sensing communication, that a critical mass of organisms has assembled, they use their flagella to slide toward each other and construct a thick, slimy biofilm." Bacterial communication, toxin production tied to intriguing cell protein, and:
"Polyphosphate is a long, chain-like molecule found in every living cell. Scientists believe that in animals, one of its roles may be to serve as a phosphate storage reservoir for the production of ATP (adenosine triphosphate), which provides the energy to power a cell. In bacteria, polyphosphate helps these single-celled organisms adapt to nutritional deficiencies and environmental stresses. It also helps them survive the state of suspended animation known as the stationary phase of growth. "
Bacteria also communicate via conjugation, which enables bacteria to exchange genetic material because of tube-like connections called pili. The donor bacterium contains conjugative or mobilizable genetic elements, usually a conjugative plasmid or episome plasmid that can integrate itself into the bacterial chromosome by genetic recombination. One such conjugative plasmid is called the F-plasmid. This is an episome about 100 thousand base-pairs in length, which carries its own origin of replication, called oriV. Most conjugative plasmids have systems ensuring that the recipient cell does not already contain a similar element, ensuring that there is only one copy of the F-plasmid in the F-positive bacterium. Genetic Construct for F and R cell types.
: New Salmonella Finding—Inter-Bacterial Communication! : Princeton - Bacterial communication :
Mahadevan, S (2002) Bacterial communication. Journal of Biosciences 27(5):pp. 443-444. Full text available as:PDF.
Phosphorylation switches
response regulator proteins in bacteria
Molecular Mechanisms of Signal Transduction : "The majority of bacterial response regulator proteins are transcription factors that serve as repressors or activators to regulate the expression of specific genes. The effector domains of these response regulators are DNA-binding domains that can be categorized into three major families based on sequence and structural similarity.
The OmpR/PhoB family of response regulator transcription factors, distinguished by a winged-helix DNA-binding domain, is the largest family, accounting for ~45 percent of all response regulators. Most characterized members of this family have been shown to bind as tandem dimers to direct repeat DNA recognition sequences. . . Phosphorylation induces dimerization or higher-order oligomerization of the proteins and that dimerization is mediated by the phosphorylated regulatory domains. The activated regulatory domains of Escherichia coli ArcA, KdpE, PhoB, PhoP, and TorR and T. maritima DrrB and DrrD all exist as dimers with identical alpha4-beta5-alpha5 interfaces. In all cases, the dimerization interface is formed by a few hydrophobic residues surrounded by an extensive network of intra- and intermolecular salt bridges. The residues involved in these interactions are highly conserved in all members of the OmpR/PhoB family, but not in other response regulators. It is proposed that this mode of dimerization is common to most members of the OmpR/PhoB family and that it represents a family-specific mechanism for activation of DNA binding. Upon phosphorylation, the interface between regulatory and DNA-binding domains is disrupted, allowing the regulatory domains to dimerize via their alpha4-beta5-aalpha5 faces. Disruption of the interdomain interface frees the DNA-binding domain, allowing it to dimerize in tandem on direct repeat DNA half-sites with symmetry that is different from that of the regulatory domain dimer. Additional nuclear magnetic resonance (NMR) and biochemical studies support this mechanism of activation. " Link to link-to-images.








































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}