Bacterial motility
Many prokaryotes are not motile, however motile prokaryotes employ a number of mechanisms:
1. Flagella - Gram -ve, Gram +ve
2. Gliding
3. Buoyancy change
4. Axial filaments - Spirochetes,
Prokaryotic movement is controlled by chemotactic signaling (below)
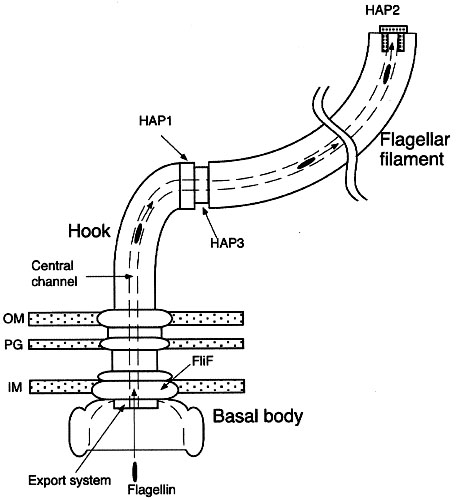
Bacterial flagella are rigid, hollow structures constructed of the protein flagellin (FliC). Each long filament is attached by a hook to the basal body, which serves as the motor.
The filament is usually about 20 nm in diameter and typically comprises of thousands of copies of a single form of flagellin. Less often the filament is composed of several different flagellins. A capping protein, HAP2, is located at the tip of the flagellum. The junction of the single-protein hook and filament requires hook-associated proteins called HAP1 and HAP3. The basal bodies of Gram +ve bacteria are anchored in the plasma membrane. The basal structure comprises a rod, a series of rings, the Mot proteins, the switch complex and the flagellum-specific export apparatus. The rings anchor the flagellum to the cytoplasmic membrane (MS ring), the peptidoglycan (P ring) and the outer membrane (L ring). The Gram -ve bacteria lack the P and L rings, and instead have an additional basal body embedded in the outer membrane. The switch proteins (FliG, FliM and FliN) permit the flagellum to switch rotation, thus controlling the direction of swimming in response to attractants or repellents in the environment (chemotaxis system). As a result of environmental sensing, phosphorylated CheY protein comes into direct contact with the FliM switch protein (Bourret et al., 2002). MotA and MotB proteins form a channel through which flow the protons that power the rotation of the flagellum. They form the stator, or nonrotating portion, of the structure where MotB is apparently attached to the peptidoglycan layer. The rotor extends into the cytoplasm (forming the C ring) and comprising several proteins including the three switch proteins. Image with legend : Diagram : image bacterial flagellar nanomotor : diagram :
Note: the bacterial flagellum is the target of intellectualized creationist nonsense in the guise of claims of so-called "irreducible complexity", which take the position that assemblages of the individual components of molecular machinery could not have evolved, but could only be explained by design (God). For refutation see: Research has demonstrated that assembly of pre-existing modifications operate in subsequently evolved features.* Reducible complexity : "As an icon of anti-evolution, the flagellum has fallen." The Flagellum unspun : Flagellum evolution in Nature Reviews Microbiology re Pallen MJ, Matzke NJ. (2006). “From The Origin of Species to the origin of bacterial flagella.” Nature Reviews Microbiology, 4(10), 784-790. October 2006. Advanced Online Publication on September 5, 2006. [PubMed] [Journal] [DOI] [Google Scholar] : Evolution in (Brownian) space: a model for the origin of the bacterial flagellum & Background to "Evolution in (Brownian) space: a model for the origin of the bacterial flagellum" : Inventing the dynamo machine: the evolution of the F-type and V-type ATPases :
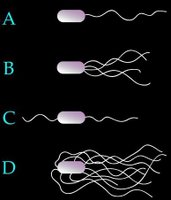 Arrangement of eubacterial flagella
Arrangement of eubacterial flagella
A. monotrichous – polar – Pseudomonas sp.,
B. lophotrichous – Spirillum
C. amphitrichous –
D. peritrichous – Proteus vulgaris, sem - peritrichous flagella, E. coli,
Mechanisms of flagellar motility in eubacteria:
Motile behavior of bacteria : Sense and sensibility in bacteria : Animated motor :
Archaeal flagella are superficially similar to those of eubacteria, but are not homologous to bacterial flagella because of a number of differences in structure, power source, and growth. [1, 2, 3, 4] Diagram - Archaeal flagellum :
The Archaeal flagellum is:
1. only 15 nm in diameter, rather than 20 nm as in eubacteria.
2. appears to grow at base rather than tip
3. is homologous to Type IV pili, unlike eubacteria
4. is coded by genes without sequence homology to the eubacteria
The archaeal flagellum: a different kind of prokaryotic motility structure.
The archaeal flagellum is a unique motility apparatus distinct in composition and likely in assembly from the bacterial flagellum. Gene families comprised of multiple flagellin genes co-transcribed with a number of conserved, archaeal-specific accessory genes have been identified in several archaea. However, no homologues of any bacterial genes involved in flagella structure have yet been identified in any archaeon, including those archaea in which the complete genome sequence has been published. Archaeal flagellins possess a highly conserved hydrophobic N-terminal sequence that is similar to that of type IV pilins and clearly unlike that of bacterial flagellins. Also unlike bacterial flagellins but similar to type IV pilins, archaeal flagellins are initially synthesized with a short leader peptide that is cleaved by a membrane-located peptidase. Thomas NA, Bardy SL, Jarrell KF. The archaeal flagellum: a different kind of prokaryotic motility structure. FEMS Microbiol Rev. 2001 Apr;25(2):147-74.
Recent advances in the structure and assembly of the archaeal flagellum. [J Mol Microbiol Biotechnol. 2004] PMID: 15170402
Characterization of flagellum gene families of methanogenic archaea and localization of novel flagellum accessory proteins. [J Bacteriol. 2001] PMID: 11717274
Prokaryotic motility structures. [Microbiology. 2003] PMID: 12624192
Mutants in flaI and flaJ of the archaeon Methanococcus voltae are deficient in flagellum assembly. [Mol Microbiol. 2002] PMID: 12410843
Stress genes and proteins in the archaea. [Microbiol Mol Biol Rev. 1999]
Both eubacterial and archaeal flagella differ from the flagella of eukaryotes:
Eukaryotic cilia and flagella both have an internal structure built upon microtubules, but the flagellum is longer and is more often a single organelle. Inside both cilia and flagella is a microtubule-based cytoskeleton termed the axoneme, which provides scaffolding for various protein complexes.
Chemotaxis is a signal transduction systems that controls movement. Even though the motility apparatus differs among organisms, the general mechanism of control of chemotaxis is conserved throughout all bacteria and archaea. The chemotaxis mechanism in B. subtilis is probably close to that of the ancestral organism from which the bacteria and archaea descended. The chemotactic system of E. coli is streamlined and lacks, or has significantly modified, some basic features of the primordial mechanism that existed when the bacteria and archaea separated during biological evolution.
Central to chemotactic control is the two-component system in which phosphorylation of a response regulator reflects phosphorylation of a histidine autokinase that senses environmental parameters (117). This is the commonest mode of signal transduction system in bacteria, and the "two component system" controls diverse processes such as gene expression, sporulation, and chemotaxis. • Phosphorylation switches •
In prokaryotes, most signaling is effected by a simple two-component systems (TCS), which may bring about changes in gene transcription or chemotactic (swimming) behavior of the organism. In bacteria, most TCS are produced by one histidine kinase (HK) and its corresponding response regulator (RR). After detection of a signal, such as an alteration in the cell's redox status or the ion concentration in the medium, two HK monomers dimerize and phosphorylate a histidine residue in trans. Subsequently the phosphate group is transferred to an aspartate residue of the RR. This short phosphorylation cascade (figure showing basic two-component phosphotransfer scheme) can impact chemotaxis (via the CheA-CheY-system) and the pattern of gene expression. At the receptors, chemotactic signals control autophosphorylation of the CheA histidine kinase. The phosphohistidine acts as a substrate for the response regulator CheY, which catalyzes the transfer of the phosphoryl group to a conserved aspartate (review 250). The resulting phosphorylated CheY-P can interact with the switch mechanism in the motor (42, 149, 186, 193, 234), causing a change in movement, such as in direction or speed of rotation of flagella.
Chemotaxis proteins comprise four groups—a signal recognition and transduction group, an excitation group, an adaptation group, and a signal removal group (to dephosphorylate CheY-P). The signal recognition and transduction group includes the receptors (9, 81, 118) and ligand binding proteins (4, 86), which are capable of binding effectors outside the cell; a few receptors, however, are cytoplasmic (92, 93, 229). Image general chemotaxis model : Table proteins in chemotaxis :
Some prokaryotic TCSs are more sophisticated than the basic HK-RR system. These comprise a "hybridkinase", which consists of a kinase domain and a response regulator domain plus an additional phosphorylatable histidine residue. These modified TCS are also called "phosphorelays". Because two or more phosphorylation events (figure showing phosphorelay system) occur, it is believed that these systems can integrate more signals into the signaling cascade and are more fine tuned. Prokaryotic phosphorelays are a minority in the TCS-family, whereas they are the major type of two-component systems found in eukaryotes. Increasingly, TCSs have been demonstrated to cross-communicate (figure showing cross talk between the Pho- and Pmr-TCS in Salmonella enterica), enhancing the organism's responsiveness to the environment.
Abstracts of Articles:
Diversity in chemotaxis mechanisms among the bacteria and archaea.
The study of chemotaxis describes the cellular processes that control the movement of organisms toward favorable environments. In bacteria and archaea, motility is controlled by a two-component system involving a histidine kinase that senses the environment and a response regulator, a very common type of signal transduction in prokaryotes. Most insights into the processes involved have come from studies of Escherichia coli over the last three decades. However, in the last 10 years, with the sequencing of many prokaryotic genomes, it has become clear that E. coli represents a streamlined example of bacterial chemotaxis. While general features of excitation remain conserved among bacteria and archaea, specific features, such as adaptational processes and hydrolysis of the intracellular signal CheY-P, are quite diverse. The Bacillus subtilis chemotaxis system is considerably more complex and appears to be similar to the one that existed when the bacteria and archaea separated during evolution, so that understanding this mechanism should provide insight into the variety of mechanisms used today by the broad sweep of chemotactic bacteria and archaea. However, processes even beyond those used in E. coli and B. subtilis have been discovered in other organisms. This review emphasizes those used by B. subtilis and these other organisms but also gives an account of the mechanism in E. coli.
Szurmant H, Ordal GW. Diversity in Chemotaxis Mechanisms among the Bacteria and Archaea. (Free Full Text Article) Microbiol Mol Biol Rev. 2004 Jun;68(2):301-19.
Prokaryotic motility structures.
Prokaryotes use a wide variety of structures to facilitate motility. The majority of research to date has focused on swimming motility and the molecular architecture of the bacterial flagellum. While intriguing questions remain, especially concerning the specialized export system involved in flagellum assembly, for the most part the structural components and their location within the flagellum and function are now known. The same cannot be said of the other apparati including archaeal flagella, type IV pili, the junctional pore, ratchet structure and the contractile cytoskeleton used by a variety of organisms for motility. In these cases, many of the structural components have yet to be identified and the mechanism of action that results in motility is often still poorly understood. Research on the bacterial flagellum has greatly aided our understanding of not only motility but also protein secretion and genetic regulation systems. Continued study and understanding of all prokaryotic motility structures will provide a wealth of knowledge that is sure to extend beyond the bounds of prokaryotic movement.
Bardy SL, Ng SY, Jarrell KF. Prokaryotic motility structures. (Free Full Text Article) Microbiology. 2003 Feb;149(Pt 2):295-304.
The archaeal flagellum: a different kind of prokaryotic motility structure. [FEMS Microbiol Rev. 2001] PMID: 11250034
Recent advances in the structure and assembly of the archaeal flagellum. [J Mol Microbiol Biotechnol. 2004] PMID: 15170402
Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. [Microbiology. 2003] PMID: 14600218
Diversity in chemotaxis mechanisms among the bacteria and archaea. [Microbiol Mol Biol Rev. 2004] PMID: 15187186
Bacterial and archaeal flagella as prokaryotic motility organelles. [Biochemistry (Mosc). 2004] PMID: 15627373
See all Related Articles...
Characterization of flagellum gene families of methanogenic archaea and localization of novel flagellum accessory proteins. (Free Full Text Article) Thomas NA, Jarrell KF. J Bacteriol. 2001 Dec;183(24):7154-64.
Bourret,R. B., Charon, N. W., Stock, A. M. & West, A. H. (2002). Bright lights, abundant operons – fluorescence and genomic technologies advance studies of bacterial locomotion and signal transduction: review of the BLAST meeting, Cuernavaca, Mexico, 14–19 January 2001. J Bacteriol 184, 1–17. [Free Full Text]
3D diagram - axoneme : 3D animation – inside flagellum : image - detail of cilia : tem - structure cilium : diagram - mechanism of ciliary motility : Geometric Clutch Model : animation - cilia & flagella
1. Flagella - Gram -ve, Gram +ve
2. Gliding
3. Buoyancy change
4. Axial filaments - Spirochetes,
Prokaryotic movement is controlled by chemotactic signaling (below)
Bacterial flagella are rigid, hollow structures constructed of the protein flagellin (FliC). Each long filament is attached by a hook to the basal body, which serves as the motor.
The filament is usually about 20 nm in diameter and typically comprises of thousands of copies of a single form of flagellin. Less often the filament is composed of several different flagellins. A capping protein, HAP2, is located at the tip of the flagellum. The junction of the single-protein hook and filament requires hook-associated proteins called HAP1 and HAP3. The basal bodies of Gram +ve bacteria are anchored in the plasma membrane. The basal structure comprises a rod, a series of rings, the Mot proteins, the switch complex and the flagellum-specific export apparatus. The rings anchor the flagellum to the cytoplasmic membrane (MS ring), the peptidoglycan (P ring) and the outer membrane (L ring). The Gram -ve bacteria lack the P and L rings, and instead have an additional basal body embedded in the outer membrane. The switch proteins (FliG, FliM and FliN) permit the flagellum to switch rotation, thus controlling the direction of swimming in response to attractants or repellents in the environment (chemotaxis system). As a result of environmental sensing, phosphorylated CheY protein comes into direct contact with the FliM switch protein (Bourret et al., 2002). MotA and MotB proteins form a channel through which flow the protons that power the rotation of the flagellum. They form the stator, or nonrotating portion, of the structure where MotB is apparently attached to the peptidoglycan layer. The rotor extends into the cytoplasm (forming the C ring) and comprising several proteins including the three switch proteins. Image with legend : Diagram : image bacterial flagellar nanomotor : diagram :
Note: the bacterial flagellum is the target of intellectualized creationist nonsense in the guise of claims of so-called "irreducible complexity", which take the position that assemblages of the individual components of molecular machinery could not have evolved, but could only be explained by design (God). For refutation see: Research has demonstrated that assembly of pre-existing modifications operate in subsequently evolved features.* Reducible complexity : "As an icon of anti-evolution, the flagellum has fallen." The Flagellum unspun : Flagellum evolution in Nature Reviews Microbiology re Pallen MJ, Matzke NJ. (2006). “From The Origin of Species to the origin of bacterial flagella.” Nature Reviews Microbiology, 4(10), 784-790. October 2006. Advanced Online Publication on September 5, 2006. [PubMed] [Journal] [DOI] [Google Scholar] : Evolution in (Brownian) space: a model for the origin of the bacterial flagellum & Background to "Evolution in (Brownian) space: a model for the origin of the bacterial flagellum" : Inventing the dynamo machine: the evolution of the F-type and V-type ATPases :
Arrangement of eubacterial flagellaA. monotrichous – polar – Pseudomonas sp.,
B. lophotrichous – Spirillum
C. amphitrichous –
D. peritrichous – Proteus vulgaris, sem - peritrichous flagella, E. coli,
Mechanisms of flagellar motility in eubacteria:
Motile behavior of bacteria : Sense and sensibility in bacteria : Animated motor :
Archaeal flagella are superficially similar to those of eubacteria, but are not homologous to bacterial flagella because of a number of differences in structure, power source, and growth. [1, 2, 3, 4] Diagram - Archaeal flagellum :
The Archaeal flagellum is:
1. only 15 nm in diameter, rather than 20 nm as in eubacteria.
2. appears to grow at base rather than tip
3. is homologous to Type IV pili, unlike eubacteria
4. is coded by genes without sequence homology to the eubacteria
The archaeal flagellum: a different kind of prokaryotic motility structure.
The archaeal flagellum is a unique motility apparatus distinct in composition and likely in assembly from the bacterial flagellum. Gene families comprised of multiple flagellin genes co-transcribed with a number of conserved, archaeal-specific accessory genes have been identified in several archaea. However, no homologues of any bacterial genes involved in flagella structure have yet been identified in any archaeon, including those archaea in which the complete genome sequence has been published. Archaeal flagellins possess a highly conserved hydrophobic N-terminal sequence that is similar to that of type IV pilins and clearly unlike that of bacterial flagellins. Also unlike bacterial flagellins but similar to type IV pilins, archaeal flagellins are initially synthesized with a short leader peptide that is cleaved by a membrane-located peptidase. Thomas NA, Bardy SL, Jarrell KF. The archaeal flagellum: a different kind of prokaryotic motility structure. FEMS Microbiol Rev. 2001 Apr;25(2):147-74.
Recent advances in the structure and assembly of the archaeal flagellum. [J Mol Microbiol Biotechnol. 2004] PMID: 15170402
Characterization of flagellum gene families of methanogenic archaea and localization of novel flagellum accessory proteins. [J Bacteriol. 2001] PMID: 11717274
Prokaryotic motility structures. [Microbiology. 2003] PMID: 12624192
Mutants in flaI and flaJ of the archaeon Methanococcus voltae are deficient in flagellum assembly. [Mol Microbiol. 2002] PMID: 12410843
Stress genes and proteins in the archaea. [Microbiol Mol Biol Rev. 1999]
Both eubacterial and archaeal flagella differ from the flagella of eukaryotes:
Eukaryotic cilia and flagella both have an internal structure built upon microtubules, but the flagellum is longer and is more often a single organelle. Inside both cilia and flagella is a microtubule-based cytoskeleton termed the axoneme, which provides scaffolding for various protein complexes.
Chemotaxis is a signal transduction systems that controls movement. Even though the motility apparatus differs among organisms, the general mechanism of control of chemotaxis is conserved throughout all bacteria and archaea. The chemotaxis mechanism in B. subtilis is probably close to that of the ancestral organism from which the bacteria and archaea descended. The chemotactic system of E. coli is streamlined and lacks, or has significantly modified, some basic features of the primordial mechanism that existed when the bacteria and archaea separated during biological evolution.
Central to chemotactic control is the two-component system in which phosphorylation of a response regulator reflects phosphorylation of a histidine autokinase that senses environmental parameters (117). This is the commonest mode of signal transduction system in bacteria, and the "two component system" controls diverse processes such as gene expression, sporulation, and chemotaxis. • Phosphorylation switches •
In prokaryotes, most signaling is effected by a simple two-component systems (TCS), which may bring about changes in gene transcription or chemotactic (swimming) behavior of the organism. In bacteria, most TCS are produced by one histidine kinase (HK) and its corresponding response regulator (RR). After detection of a signal, such as an alteration in the cell's redox status or the ion concentration in the medium, two HK monomers dimerize and phosphorylate a histidine residue in trans. Subsequently the phosphate group is transferred to an aspartate residue of the RR. This short phosphorylation cascade (figure showing basic two-component phosphotransfer scheme) can impact chemotaxis (via the CheA-CheY-system) and the pattern of gene expression. At the receptors, chemotactic signals control autophosphorylation of the CheA histidine kinase. The phosphohistidine acts as a substrate for the response regulator CheY, which catalyzes the transfer of the phosphoryl group to a conserved aspartate (review 250). The resulting phosphorylated CheY-P can interact with the switch mechanism in the motor (42, 149, 186, 193, 234), causing a change in movement, such as in direction or speed of rotation of flagella.
Chemotaxis proteins comprise four groups—a signal recognition and transduction group, an excitation group, an adaptation group, and a signal removal group (to dephosphorylate CheY-P). The signal recognition and transduction group includes the receptors (9, 81, 118) and ligand binding proteins (4, 86), which are capable of binding effectors outside the cell; a few receptors, however, are cytoplasmic (92, 93, 229). Image general chemotaxis model : Table proteins in chemotaxis :
Some prokaryotic TCSs are more sophisticated than the basic HK-RR system. These comprise a "hybridkinase", which consists of a kinase domain and a response regulator domain plus an additional phosphorylatable histidine residue. These modified TCS are also called "phosphorelays". Because two or more phosphorylation events (figure showing phosphorelay system) occur, it is believed that these systems can integrate more signals into the signaling cascade and are more fine tuned. Prokaryotic phosphorelays are a minority in the TCS-family, whereas they are the major type of two-component systems found in eukaryotes. Increasingly, TCSs have been demonstrated to cross-communicate (figure showing cross talk between the Pho- and Pmr-TCS in Salmonella enterica), enhancing the organism's responsiveness to the environment.
Abstracts of Articles:
Diversity in chemotaxis mechanisms among the bacteria and archaea.
The study of chemotaxis describes the cellular processes that control the movement of organisms toward favorable environments. In bacteria and archaea, motility is controlled by a two-component system involving a histidine kinase that senses the environment and a response regulator, a very common type of signal transduction in prokaryotes. Most insights into the processes involved have come from studies of Escherichia coli over the last three decades. However, in the last 10 years, with the sequencing of many prokaryotic genomes, it has become clear that E. coli represents a streamlined example of bacterial chemotaxis. While general features of excitation remain conserved among bacteria and archaea, specific features, such as adaptational processes and hydrolysis of the intracellular signal CheY-P, are quite diverse. The Bacillus subtilis chemotaxis system is considerably more complex and appears to be similar to the one that existed when the bacteria and archaea separated during evolution, so that understanding this mechanism should provide insight into the variety of mechanisms used today by the broad sweep of chemotactic bacteria and archaea. However, processes even beyond those used in E. coli and B. subtilis have been discovered in other organisms. This review emphasizes those used by B. subtilis and these other organisms but also gives an account of the mechanism in E. coli.
Szurmant H, Ordal GW. Diversity in Chemotaxis Mechanisms among the Bacteria and Archaea. (Free Full Text Article) Microbiol Mol Biol Rev. 2004 Jun;68(2):301-19.
Prokaryotic motility structures.
Prokaryotes use a wide variety of structures to facilitate motility. The majority of research to date has focused on swimming motility and the molecular architecture of the bacterial flagellum. While intriguing questions remain, especially concerning the specialized export system involved in flagellum assembly, for the most part the structural components and their location within the flagellum and function are now known. The same cannot be said of the other apparati including archaeal flagella, type IV pili, the junctional pore, ratchet structure and the contractile cytoskeleton used by a variety of organisms for motility. In these cases, many of the structural components have yet to be identified and the mechanism of action that results in motility is often still poorly understood. Research on the bacterial flagellum has greatly aided our understanding of not only motility but also protein secretion and genetic regulation systems. Continued study and understanding of all prokaryotic motility structures will provide a wealth of knowledge that is sure to extend beyond the bounds of prokaryotic movement.
Bardy SL, Ng SY, Jarrell KF. Prokaryotic motility structures. (Free Full Text Article) Microbiology. 2003 Feb;149(Pt 2):295-304.
The archaeal flagellum: a different kind of prokaryotic motility structure. [FEMS Microbiol Rev. 2001] PMID: 11250034
Recent advances in the structure and assembly of the archaeal flagellum. [J Mol Microbiol Biotechnol. 2004] PMID: 15170402
Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. [Microbiology. 2003] PMID: 14600218
Diversity in chemotaxis mechanisms among the bacteria and archaea. [Microbiol Mol Biol Rev. 2004] PMID: 15187186
Bacterial and archaeal flagella as prokaryotic motility organelles. [Biochemistry (Mosc). 2004] PMID: 15627373
See all Related Articles...
Characterization of flagellum gene families of methanogenic archaea and localization of novel flagellum accessory proteins. (Free Full Text Article) Thomas NA, Jarrell KF. J Bacteriol. 2001 Dec;183(24):7154-64.
Bourret,R. B., Charon, N. W., Stock, A. M. & West, A. H. (2002). Bright lights, abundant operons – fluorescence and genomic technologies advance studies of bacterial locomotion and signal transduction: review of the BLAST meeting, Cuernavaca, Mexico, 14–19 January 2001. J Bacteriol 184, 1–17. [Free Full Text]
3D diagram - axoneme : 3D animation – inside flagellum : image - detail of cilia : tem - structure cilium : diagram - mechanism of ciliary motility : Geometric Clutch Model : animation - cilia & flagella
Labels: Archaea, bacterial flagellum, bacterial motility, chemotactic signaling, chemotaxis, Gram stain, kinase, peptidoglycan, two-componets systems








































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
0 Glossary:
Post a Comment
<< Home