Stress genes, proteins, molecular chaperones
Activation of stress genes is an important component of the stress response (90, 120, 205, 208, 219, 253, 254, 279)[s]. Stress (physical or chemical) also inactivates or down-regulates many genes, including many housekeeping genes.
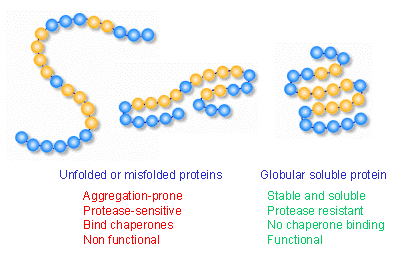
Stress proteins, which are encoded by stress genes, play critical roles in physiological protein biogenesis. These proteins assist in the folding, translocation, and assembly of other proteins (191, 214, 235, 236, 247), so many stress proteins are also called molecular chaperones (72). Not all stress proteins are chaperones and, conversely, not every molecular chaperone is a stress protein. : image 24 unit Archaebacterial heat shock protein :
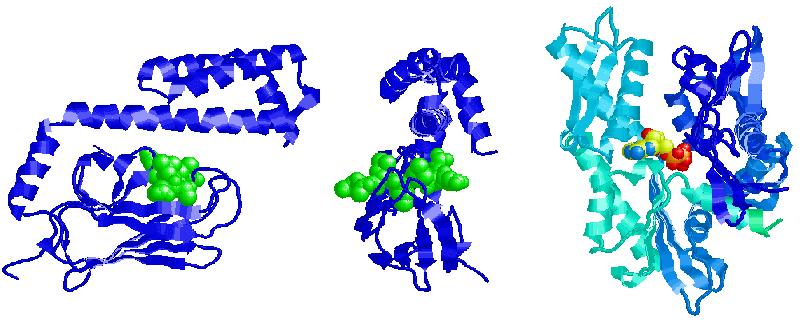
Molecular chaperones interact with unfolded or partially folded protein subunits, such as nascent chains emerging from the ribosome or extended chains undergoing translocation across subcellular membranes. The chief function of chaperones is to prevent inappropriate association or aggregation of exposed hydrophobic surfaces by directing their substrates into productive folding, transport or degradation pathways. Chaperones stabilize non-native conformation and facilitate correct folding of protein subunits, often coupling ATP binding/hydrolysis to the folding process. They do not interact with native proteins, nor do they form part of the final folded structures. Some chaperones are non-specific in that they interact with a wide variety of polypeptide chains, while other chaperoness are restricted to specific targets. Chaperones are essential for viability, so their expression is often increased by cellular stress. : image substrate binding by chaperones : image Hsp60 chaperonin GroEL : image Hsp70 : more :
Stress proteins help other cellular proteins to (i) fold correctly during and after translation; (ii) migrate to the cell's locale, where they will reside and function; and (iii) assemble into the quaternary structure that will make them useful to the cell when the proteins function as polymers. Further, some stress proteins participate in the degradation of other polypeptides such as those that are denatured beyond recovery and could pose a serious threat to the cell if they were to aggregate (39, 92, 97, 98, 122, 123, 133).
Thermophilic cell lines adapted for survival at high temperatures constitutively synthesize the stress protein hsp70 at high levels, whereas temperature-sensitive cell-lines do not. Several stress protein families, including hsp90, hsp70, chaperonin60, hsp40, the low molecular weight stress proteins, and ubiquitin, are expressed in diverse phyla. Studies comparing the stress protein repetoir of closely related species inhabiting different environments have demonstrated that differences in the heat-stress response are correlated with thermal resistance, suggesting that stress proteins help organisms adapt to harsh or unpredictable environments.
. cellular stress response .
Stress genes and proteins in the archaea.
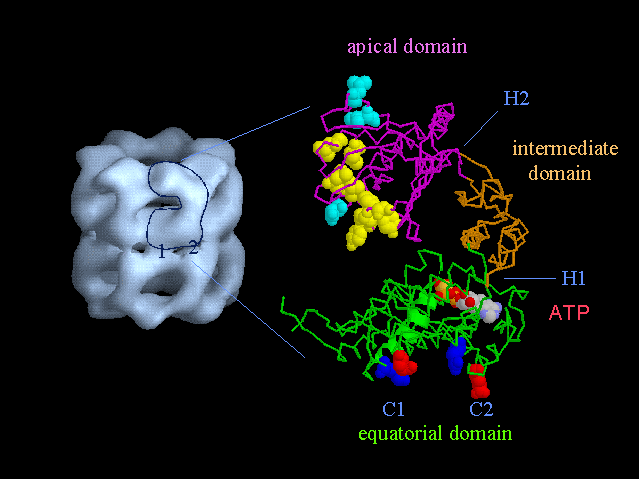
The field covered in this review is new; the first sequence of a gene encoding the molecular chaperone Hsp70 and the first description of a chaperonin in the archaea were reported in 1991. These findings boosted research in other areas beyond the archaea that were directly relevant to bacteria and eukaryotes, for example, stress gene regulation, the structure-function relationship of the chaperonin complex, protein-based molecular phylogeny of organisms and eukaryotic-cell organelles, molecular biology and biochemistry of life in extreme environments, and stress tolerance at the cellular and molecular levels. In the last 8 years, archaeal stress genes and proteins belonging to the families Hsp70, Hsp60 (chaperonins), Hsp40(DnaJ), and small heat-shock proteins (sHsp) have been studied. The hsp70(dnaK), hsp40(dnaJ), and grpE genes (the chaperone machine) have been sequenced in seven, four, and two species, respectively, but their expression has been examined in detail only in the mesophilic methanogen Methanosarcina mazei S-6. The proteins possess markers typical of bacterial homologs but none of the signatures distinctive of eukaryotes. In contrast, gene expression and transcription initiation signals and factors are of the eucaryal type, which suggests a hybrid archaeal-bacterial complexion for the Hsp70 system. Another remarkable feature is that several archaeal species in different phylogenetic branches do not have the gene hsp70(dnaK), an evolutionary puzzle that raises the important question of what replaces the product of this gene, Hsp70(DnaK), in protein biogenesis and refolding and for stress resistance. Although archaea are prokaryotes like bacteria, their Hsp60 (chaperonin) family is of type (group) II, similar to that of the eukaryotic cytosol; however, unlike the latter, which has several different members, the archaeal chaperonin system usually includes only two (in some species one and in others possibly three) related subunits of approximately 60 kDa. These form, in various combinations depending on the species, a large structure or chaperonin complex sometimes called the thermosome. This multimolecular assembly is similar to the bacterial chaperonin complex GroEL/S, but it is made of only the large, double-ring oligomers each with eight (or nine) subunits instead of seven as in the bacterial complex. Like Hsp70(DnaK), the archaeal chaperonin subunits are remarkable for their evolution, but for a different reason. Ubiquitous among archaea, the chaperonins show a pattern of recurrent gene duplication-hetero-oligomeric chaperonin complexes appear to have evolved several times independently. The stress response and stress tolerance in the archaea involve chaperones, chaperonins, other heat shock (stress) proteins including sHsp, thermoprotectants, the proteasome, as yet incompletely understood thermoresistant features of many molecules, and formation of multicellular structures. The latter structures include single- and mixed-species (bacterial-archaeal) types. Many questions remain unanswered, and the field offers extraordinary opportunities owing to the diversity, genetic makeup, and phylogenetic position of archaea and the variety of ecosystems they inhabit. Specific aspects that deserve investigation are elucidation of the mechanism of action of the chaperonin complex at different temperatures, identification of the partners and substitutes for the Hsp70 chaperone machine, analysis of protein folding and refolding in hyperthermophiles, and determination of the molecular mechanisms involved in stress gene regulation in archaeal species that thrive under widely different conditions (temperature, pH, osmolarity, and barometric pressure). These studies are now possible with uni- and multicellular archaeal models and are relevant to various areas of basic and applied research, including exploration and conquest of ecosystems inhospitable to humans and many mammals and plants.
Macario AJ, Lange M, Ahring BK, De Macario EC. Stress genes and proteins in the archaea. (Free Full Text Article) Microbiol Mol Biol Rev. 1999 Dec;63(4):923-67, table of contents.
The archaeal molecular chaperone machine: peculiarities and paradoxes. [Genetics. 1999] PMID: 10430558
Molecular biology of stress genes in methanogens: potential for bioreactor technology. [Adv Biochem Eng Biotechnol. 2003] PMID: 12747562
The molecular chaperone system and other anti-stress mechanisms in archaea. [Front Biosci. 2001] PMID: 11171552
Evolution of assisted protein folding: the distribution of the main chaperoning systems within the phylogenetic domain archaea. [Front Biosci. 2004] PMID: 14977547
The basal transcription factors TBP and TFB from the mesophilic archaeon Methanosarcina mazeii: structure and conformational changes upon interaction with stress-gene promoters. [J Mol Biol. 2001] PMID: 11397082
See all Related Articles...
Stress proteins, which are encoded by stress genes, play critical roles in physiological protein biogenesis. These proteins assist in the folding, translocation, and assembly of other proteins (191, 214, 235, 236, 247), so many stress proteins are also called molecular chaperones (72). Not all stress proteins are chaperones and, conversely, not every molecular chaperone is a stress protein. : image 24 unit Archaebacterial heat shock protein :
Molecular chaperones interact with unfolded or partially folded protein subunits, such as nascent chains emerging from the ribosome or extended chains undergoing translocation across subcellular membranes. The chief function of chaperones is to prevent inappropriate association or aggregation of exposed hydrophobic surfaces by directing their substrates into productive folding, transport or degradation pathways. Chaperones stabilize non-native conformation and facilitate correct folding of protein subunits, often coupling ATP binding/hydrolysis to the folding process. They do not interact with native proteins, nor do they form part of the final folded structures. Some chaperones are non-specific in that they interact with a wide variety of polypeptide chains, while other chaperoness are restricted to specific targets. Chaperones are essential for viability, so their expression is often increased by cellular stress. : image substrate binding by chaperones : image Hsp60 chaperonin GroEL : image Hsp70 : more :
Stress proteins help other cellular proteins to (i) fold correctly during and after translation; (ii) migrate to the cell's locale, where they will reside and function; and (iii) assemble into the quaternary structure that will make them useful to the cell when the proteins function as polymers. Further, some stress proteins participate in the degradation of other polypeptides such as those that are denatured beyond recovery and could pose a serious threat to the cell if they were to aggregate (39, 92, 97, 98, 122, 123, 133).
Thermophilic cell lines adapted for survival at high temperatures constitutively synthesize the stress protein hsp70 at high levels, whereas temperature-sensitive cell-lines do not. Several stress protein families, including hsp90, hsp70, chaperonin60, hsp40, the low molecular weight stress proteins, and ubiquitin, are expressed in diverse phyla. Studies comparing the stress protein repetoir of closely related species inhabiting different environments have demonstrated that differences in the heat-stress response are correlated with thermal resistance, suggesting that stress proteins help organisms adapt to harsh or unpredictable environments.
. cellular stress response .
Stress genes and proteins in the archaea.
The field covered in this review is new; the first sequence of a gene encoding the molecular chaperone Hsp70 and the first description of a chaperonin in the archaea were reported in 1991. These findings boosted research in other areas beyond the archaea that were directly relevant to bacteria and eukaryotes, for example, stress gene regulation, the structure-function relationship of the chaperonin complex, protein-based molecular phylogeny of organisms and eukaryotic-cell organelles, molecular biology and biochemistry of life in extreme environments, and stress tolerance at the cellular and molecular levels. In the last 8 years, archaeal stress genes and proteins belonging to the families Hsp70, Hsp60 (chaperonins), Hsp40(DnaJ), and small heat-shock proteins (sHsp) have been studied. The hsp70(dnaK), hsp40(dnaJ), and grpE genes (the chaperone machine) have been sequenced in seven, four, and two species, respectively, but their expression has been examined in detail only in the mesophilic methanogen Methanosarcina mazei S-6. The proteins possess markers typical of bacterial homologs but none of the signatures distinctive of eukaryotes. In contrast, gene expression and transcription initiation signals and factors are of the eucaryal type, which suggests a hybrid archaeal-bacterial complexion for the Hsp70 system. Another remarkable feature is that several archaeal species in different phylogenetic branches do not have the gene hsp70(dnaK), an evolutionary puzzle that raises the important question of what replaces the product of this gene, Hsp70(DnaK), in protein biogenesis and refolding and for stress resistance. Although archaea are prokaryotes like bacteria, their Hsp60 (chaperonin) family is of type (group) II, similar to that of the eukaryotic cytosol; however, unlike the latter, which has several different members, the archaeal chaperonin system usually includes only two (in some species one and in others possibly three) related subunits of approximately 60 kDa. These form, in various combinations depending on the species, a large structure or chaperonin complex sometimes called the thermosome. This multimolecular assembly is similar to the bacterial chaperonin complex GroEL/S, but it is made of only the large, double-ring oligomers each with eight (or nine) subunits instead of seven as in the bacterial complex. Like Hsp70(DnaK), the archaeal chaperonin subunits are remarkable for their evolution, but for a different reason. Ubiquitous among archaea, the chaperonins show a pattern of recurrent gene duplication-hetero-oligomeric chaperonin complexes appear to have evolved several times independently. The stress response and stress tolerance in the archaea involve chaperones, chaperonins, other heat shock (stress) proteins including sHsp, thermoprotectants, the proteasome, as yet incompletely understood thermoresistant features of many molecules, and formation of multicellular structures. The latter structures include single- and mixed-species (bacterial-archaeal) types. Many questions remain unanswered, and the field offers extraordinary opportunities owing to the diversity, genetic makeup, and phylogenetic position of archaea and the variety of ecosystems they inhabit. Specific aspects that deserve investigation are elucidation of the mechanism of action of the chaperonin complex at different temperatures, identification of the partners and substitutes for the Hsp70 chaperone machine, analysis of protein folding and refolding in hyperthermophiles, and determination of the molecular mechanisms involved in stress gene regulation in archaeal species that thrive under widely different conditions (temperature, pH, osmolarity, and barometric pressure). These studies are now possible with uni- and multicellular archaeal models and are relevant to various areas of basic and applied research, including exploration and conquest of ecosystems inhospitable to humans and many mammals and plants.
Macario AJ, Lange M, Ahring BK, De Macario EC. Stress genes and proteins in the archaea. (Free Full Text Article) Microbiol Mol Biol Rev. 1999 Dec;63(4):923-67, table of contents.
The archaeal molecular chaperone machine: peculiarities and paradoxes. [Genetics. 1999] PMID: 10430558
Molecular biology of stress genes in methanogens: potential for bioreactor technology. [Adv Biochem Eng Biotechnol. 2003] PMID: 12747562
The molecular chaperone system and other anti-stress mechanisms in archaea. [Front Biosci. 2001] PMID: 11171552
Evolution of assisted protein folding: the distribution of the main chaperoning systems within the phylogenetic domain archaea. [Front Biosci. 2004] PMID: 14977547
The basal transcription factors TBP and TFB from the mesophilic archaeon Methanosarcina mazeii: structure and conformational changes upon interaction with stress-gene promoters. [J Mol Biol. 2001] PMID: 11397082
See all Related Articles...








































{kind=link}
{kind=link}
{kind=link}
{kind=link}
0 Glossary:
Post a Comment
<< Home